Inicio  ÓRGANO INTERNO DE CONTROL Ciencia hoy ¿Qué nos dicen los ácidos grasos del tejido adiposo de las ballenas jorobadas?
ÓRGANO INTERNO DE CONTROL Ciencia hoy ¿Qué nos dicen los ácidos grasos del tejido adiposo de las ballenas jorobadas?
¿Qué nos dicen los ácidos grasos del tejido adiposo de las ballenas jorobadas?
Janet Nolasco Soto1 y Luis Medrano González2

Artículo publicado en la Crónica y el Portal Comunicación Veracruzana el 27 de junio 2022
¿Sabías que en los mares de México se pueden observar a ocho de las 14 especies de ballenas barbadas conocidas en el mundo? Una de ellas es la ballena jorobada y comentaremos ¿cómo? un análisis del tejido adiposo de biopsias nos permite conocer aspectos de su ecología alimentaria, estado general de sus reservas de energía y biología reproductiva.
Palabras clave: ácidos grasos, ballena jorobada, Isla Socorro.
La ballena jorobada o rorcual jorobado, Megaptera novaeangliae, es un mamífero acuático o cetáceo que se caracteriza por presentar aletas pectorales, que miden cerca de 5 m de su longitud total (13-18 m) (Figura 1). Tiene una aleta dorsal variable en aspecto que se sitúa sobre una joroba, de la cual deriva su nombre, y una aleta caudal, que ventralmente tiene una pigmentación parcial o completamente blanca con cicatrices (Figura 2). Las variaciones del color y las cicatrices en la aleta dorsal y caudal se utilizan para identificar a cada ballena jorobada pues son como el equivalente a nuestra huella digital. Posee unas láminas elásticas o barbas constituidas de queratina (como el material del que están hechas nuestras uñas), que utilizan para filtrar el alimento y que las distingue de otros cetáceos como los delfines y los cachalotes que tienen dientes. Se alimenta de microorganismos que forman el zooplancton, de krill (similar a un camarón pequeño) y de cardúmenes de peces. Tiene una capa de grasa (por debajo de su piel) que la aísla del frío. Se ha sugerido que la composición de ácidos grados del tejido adiposo de las ballenas refleja su dieta pues tiene una composición similar a la de sus presas.

A y D aletas caudales, B y C aletas dorsales de distintos individuos de ballena jorobada.
Foto: Janet Nolasco Soto
Las ballenas jorobadas deben almacenar suficientes reservas de energía en forma de grasa para realizar sus actividades reproductoras (por ej. parir a sus crías, amamantarlas, aparearse). Esto lo logran permaneciendo en aguas frías alimentándose intensamente durante el verano (cuatro a seis meses y medio) y migrando, posteriormente, en invierno hacia aguas templadas en el trópico y subtrópico de ambos hemisferios para reproducirse (Figura 3). Un aspecto importante de su ciclo migratorio es que no todas migran al mismo tiempo hacia sus áreas de alimentación o de reproducción. Esto ocurre según su edad y estado reproductivo, por ejemplo; las hembras preñadas son las primeras en llegar a las zonas de alimentación, seguidas por hembras y machos inmaduros, animales maduros de ambos sexos y al final las hembras lactantes con sus crías. Estas últimas migraran primero en el siguiente año a las zonas de reproducción seguidas por machos y hembras inmaduros, machos maduros, hembras sin conducta reproductora (anestro) y al final las hembras preñadas.
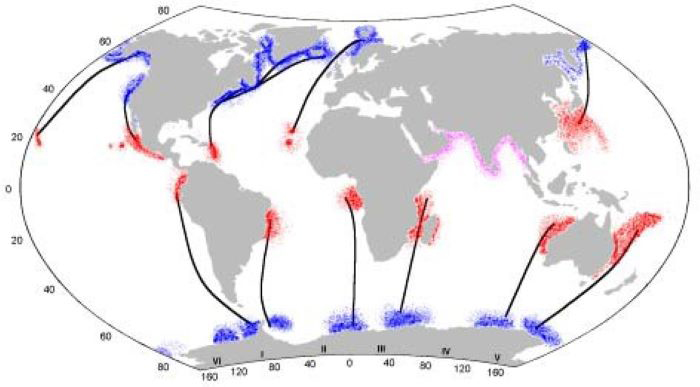
Distribución de las ballenas jorobadas en el mundo. En negro se indican las conexiones migratorias de las zonas de alimentación en verano (azul)
con las zonas de reproducción invernales (rojo). Fuente: http://www.conabio.gob.mx/institucion/proyectos/resultados/InfW024.pdf
La secuencia migratoria de las ballenas jorobadas parece relacionarse con: la capacidad para almacenar energía, la condición nutricional del individuo, así como por la abundancia y la calidad del alimento, entre otros factores. Por lo tanto, las ballenas deben hacer un uso eficiente de sus reservas de energía en el invierno durante el cual la alimentación es ocasional o nula. Por ejemplo, es sabido que las hembras preñadas de la ballena de aleta, Balaenoptera physallus, adquieren el mayor grosor de la capa de grasa durante la alimentación, en comparación con otros estadios reproductivos como machos y hembras inmaduras. Esto porque la reproducción de las hembras depende con cierto grado del almacenamiento de grasas, por lo que una disminución de presas puede reducir el número de ovulaciones y la posterior preñez. También se puede afectar la viabilidad del feto o supervivencia de la cría, disminuyendo el número de nacimientos y prolongando los intervalos de reproducción de las hembras (una cría cada dos a tres años) mientras almacenan grasas en cantidad suficiente.
Tradicionalmente, la determinación formal del estado de las reservas de energía requiere de exámenes de los animales completos, con esto en mente, exploramos las inferencias que es posible hacer a través del análisis de pequeñas muestras. En nuestro estudio analizamos la composición y concentración de ácidos grasos (contenido energético) en el tejido adiposo de biopsias de ballenas jorobadas que invernan en las aguas adyacentes a la isla Socorro, Revillagigedo, México. Esto se hizo por sexo y estado reproductivo (machos, hembras con cría, crías, hembras en anestro) en los inviernos de 1998 y 1999. Comparamos nuestros datos con la composición de ácidos grasos de ballenas jorobadas del Atlántico Norte obtenidos por Borobia y colaboradores (1995) durante el verano de 1991.
La composición de ácidos grasos fue diferente entre inviernos (1998 vs. 1999) y entre cuencas oceánicas (Pacífico Norte vs. Atlántico Norte). Por ejemplo, en las muestras de 1998 se observa mayor diversidad y concentración de ácidos grasos, que en las muestras de tejido adiposo de 1999. En la Figura 4 se puede notar la ausencia de algunos ácidos grasos en las muestras de 1999 y la presencia de otros en las muestras de 1998, los colores obscuros en las columnas indican ácidos grasos con mayor contenido energético. Esto sugiere cambios en la dieta entre inviernos. Estos cambios pueden relacionarse con el evento climático de El Niño en el verano de 1998, que fue el verano en el que se alimentaron las ballenas jorobadas que invernaron en 1999 en la isla Socorro. Este fenómeno incrementa la temperatura del mar en el Pacífico ocasionando una reducción en la productividad biológica en el océano y por tanto en las presas potenciales para las ballenas y otros animales marinos.
La diferencia entre cuencas oceánicas indica que hay una mayor variabilidad y disponibilidad de presas en el Atlántico Norte, al menos en el Golfo de San Lorenzo. Esto se nota por la presencia de más ácidos grasos en gran parte de las muestras del Atlántico Norte, así como por su mayor concentración (Figura 4). De forma global los resultados entre inviernos, sexos y estado reproductivo sugieren una marcada heterogeneidad en la alimentación de las ballenas jorobadas. Esto puede explicarse por las diferencias en comer ciertas presas de acuerdo con el sexo y/o estado reproductivo, así como por la capacidad de atraparlas, de dispersarse entre zonas de alimentación o por la disponibilidad de presas por región. Aunque, las ballenas jorobadas regresan a sus áreas tradicionales de alimentación en verano (filopatria) es posible que cambien su distribución pues se han observado algunos intercambios entre éstas en el Pacífico Norte.

Composición de ácidos grasos de la grasa de biopsias de ballenas jorobadas durante el verano de 1991 en el Atlántico Norte (Borobia et al., 1995)
y de la isla Socorro en los inviernos de 1998 y 1999. Cada columna indica un ácido graso. Los colores de la primera columna
indican a las crías (azul claro), machos (azul oscuro), hembras en anestro (amarillo) y hembras con cría (rojo).
El color en negro indica ácidos grasos más concentrados que los que tienden a colores grises claros. Crédito: Janet Nolasco Soto.
A pesar de que la cantidad de ácidos grasos en el tejido adiposo de biopsias de la ballena jorobada no refleja el estado global de las reservas de energía porque su composición y concentración varían en el grosor de la capa de grasa, entre clases de sexo/edad y entre distintas partes del cuerpo. No obstante, el análisis fisicoquímico de pequeñas muestras de tejido adiposo puede dar información sobre variaciones en la alimentación y estado general de sus reservas de energía.
Slider: Hembra con cría de ballena jorobada. Foto: Ben Phillips. https://www.pexels.com/es-es/foto/nadando-submarino-juntos-ternero-4781938/
Referencias
- Borobia M., Gearing P.G., Simard Y., Gearing J.N y Béland P. 1995. Blubber fatty acids of finback and humpback whales from the Gulf of San Lorenzo. Mar. Biol. 122: 341-353.
- Nolasco-Soto J. 2003. Contenido calórico y composición de ácidos grasos de la ballena jorobada, Megaptera novaeangliae, en la Isla Socorro, Revillagigedo, México. Tesis de Maestría, Facultad de Ciencias, UNAM. México, D.F.
1Red de Biología Evolutiva, INECOL.
2Biología Evolutiva, Facultad de Ciencias, UNAM.
